Introduction
With economic development, people’s lifestyles have gradually changed, and the number of overweight and obese individuals has increased annually. Since the 1970s, obesity has become
prevalent in developed countries, and the number of obese individuals worldwide has increased more than six times in the past
40 years, totaling more than 600 million individuals. Since 1975
and in the next 39 years, the global obesity rate (body mass index ≥30 kg/m2) increased from 3.2% to 10.8% and from 6.4% to
14.9% in women. If the trend continues, it is expected that, by
2025, 18% of men and 21% of women worldwide will be obese [1]. Overweight and obesity are not only important risk factors for
metabolic and cardiovascular diseases but are also significantly
associated with the risk of certain types of cancer (e.g., liver, kidney, prostate, colon, ovarian, breast), depression, and Alzheimer’s
disease. Moreover, the death and disability caused by obesity significantly increase the national economic burden, and the problems it poses have become a major challenge worldwide [2,3].
Being overweight and obese are equally important in child and
adolescent populations. In the past 30 years, the global situation
of obesity in children and adolescents has become critical, with
obesity detection rates increasing in many European and American countries and some Asian countries [1]. Overweight and obesity are mainly due to the fact that energy intake is greater than
consumption, energy is stored in the body in the form of fat, and
many factors can affect the body’s energy metabolism, among
which genes are a well-known cause.
Uncoupling proteins (UCPs) are thought to be involved in regulating glucose/lipid metabolism and energy homeostasis processes and are essentially a group of mitochondrial anion carrier proteins located in the inner mitochondrial membrane. As a candidate gene for obesity, Fleury et al. were the first to investigate the
UCP2 gene. The Homo sapiens UCP2 gene is located on chromosome 11q13 and consists of 8 exons and 7 introns, with a length of
8174. UCP2 is widely expressed in human tissues and is an uncoupling agent for oxidative phosphorylation, which is involved in the
regulation of lipid metabolism and ATP production, and disruption
of UCP2 expression in the adipose tissue can lead to overweight
and obesity [4,5]. UCP2 gene polymorphisms (Aal55Val, 45bp insertion/deletion, and -866G/A) have been extensively studied in
relation to overweight and obesity, but showed conflicting results.
Among these, information on the biological effects of UCP2 45bp
Ins/Del gene polymorphism is still limited. Therefore, this metaanalysis aimed to assess the effects of UCP2 45bp Ins/Del gene
polymorphism on the risk of overweight and obesity.
Methods
Search strategy
The following three databases were searched: PubMed, Embase, and China National Knowledge Infrastructure (CNKI). The
search terms included “uncoupling protein 2,” “UCP2,” “polymorphism,” “genotype,” “alleles,” “obesity,” “overweight,” and “BMI.”
The deadline for the search was September 3, 2021, for Chinese
and English literature published as electronic papers, and reference back was used to find additional literature for inclusion in
the study.
Inclusion and exclusion Criteria
Studies meeting the following criteria were included in the
meta-analysis: (1) case-control, cohort, or cross-sectional studies
assessed the association between UCP2 45bp Ins/Del gene polymorphism and overweight and obesity; (2) sufficient information
was provided in the text to calculate the frequency distribution of
the genotype of interest or that of the allele of interest; (3) subjects were recruited from the same race during the same period;
(4) if data were reported repeatedly, the most complete datasets were included; and (5) the genotype distribution of controls
obeyed the Hardy–Weinberg equilibrium. Duplicate literature, reviews, abstracts, correspondence, animal studies, non-case-control studies, and non-UCP2 45bp Ins/Del gene polymorphisms in
relation to genetic susceptibility to overweight and obesity were
excluded from this study.
Quality assessment of included studies
The Newcastle-Ottawa Scale was used to assess the quality of
each included study. Scores ≥ 7 were considered high quality.
Data extraction
The articles were evaluated, and data were independently extracted by two authors. Disagreements were resolved by consensus through discussion. The information extracted included the
following in order: name of the first author, year of publication,
country and ethnicity, number of cases and controls, frequency of
cases and control genotypes, and whether the study population
consisted of children and adolescents.
Statistical analysis
The Hardy–Weinberg equilibrium of the UCP2 45bp Ins/Del
genotype distribution in the control group was tested using the
Pearson chi-square test, and a P-value < 0.05 was considered to
deviate from the Hardy–Weinberg equilibrium. Literature deviating from the Hardy–Weinberg equilibrium was excluded from the
study. A combined odds ratio (OR) with 95% confidence interval
(CI) was calculated to assess the association between the UCP2
45bp Ins/Del gene polymorphism and the risk of genetic susceptibility to overweight and obesity in five models (allelic, dominant,
recessive, heterozygote, and homozygote models). The significance of the combined OR was determined using the Z-test. l2 was
used to assess the heterogeneity between studies. When a random-effects model was used as the combined method, heterogeneity was considered significant if l2﹥50%; otherwise, heterogeneity was considered not significantly different, and a fixed-effects
model was used. Subgroup analyses were performed for race in
each of the five genetic models. A Begg’s funnel plot was used to
identify publication bias. All statistical analyses were performed
using Stata 16.0 (StataCorp LLC, College Station, TX, USA). P-values <0.05 were considered statistically significant (two-tailed).
Results
Search results
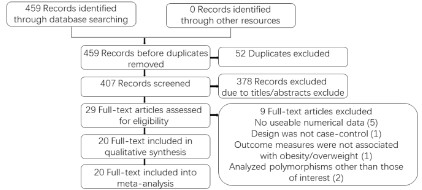
A total of 459 articles were obtained by searching PubMed,
Embase, and CNKI databases. After excluding 52 duplicate articles, 407 articles were obtained. A total of 378 articles that were
reviews, letters, laboratory studies, or not related to the current
study were excluded. Based on the abovementioned inclusion and
exclusion criteria, nine full-text articles were excluded. Twenty eligible articles were included in the meta-analysis [6-25] (Figure 1).
Study demographics
A total of 20 relevant papers were included in the meta-analysis, covering 12 countries, including 5750 cases and 6474 controls,
and of these 20 papers, five were studies on Asian populations,
and 15 were studies on Caucasian populations, with studies published between 1999 and 2020 (Table 1).
Meta-analysis
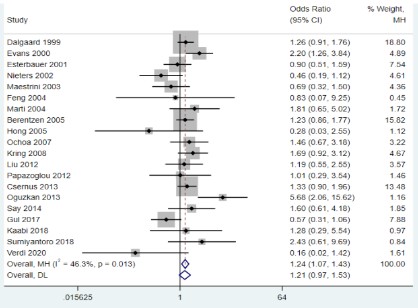
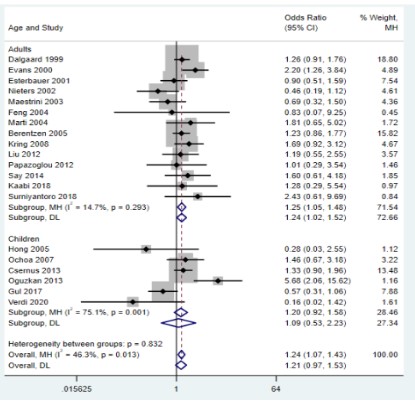
The results of the meta-analysis showed that the UCP2 45bp Ins/Del gene polymorphism was significantly associated with
genetic susceptibility to overweight and obesity only in the recessive model (OR=1.24, 95%CI=1.07-1.43, P=0.004; Figure 2),
while the other models did not show an association (P﹥0.05).
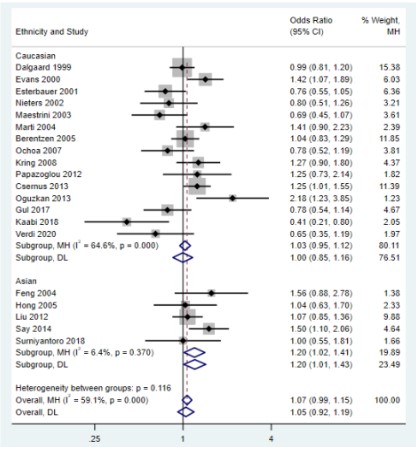
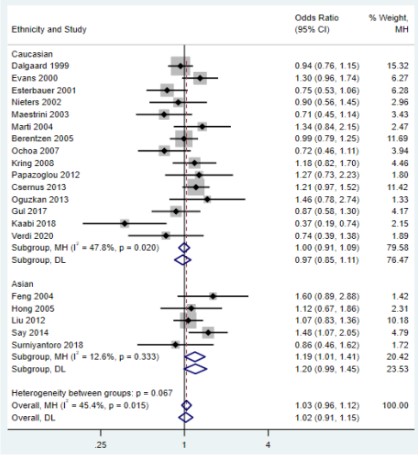
However, the subgroup analysis of ethnicity showed that, in
Asian populations, UCP2 45bp Ins/Del gene polymorphism was
allelic (OR=1.18, 95%CI=1.02-1.36, P=0.027; Figure 3), dominant
(OR=1.20, 95%CI=1.02-1.41, P=0.030; Figure 4), and heterozygote (OR=1.19, 95%CI=1.01-1.41, P=0.043; Figure 5), respectively,
which were significantly associated with genetic susceptibility to
overweight and obesity in the model. No significant associations
were found in the Caucasian population (P﹥0.05), and there was
significant heterogeneity between studies. We also performed
subgroup analysis according to whether the subjects were adults
and found significant associations only in the recessive (OR=1.19,
95% CI=1.01-1.41, P=0.011; Figure 6) and homozygote (OR=1.24, 95% CI=1.04-1.48, P=0.017; Figure 7) models in the adult group,
while no significant associations were found in the other groups
(P﹥0.05) (Table 2).
Influence analysis
To assess the stability of the study results, we performed sensitivity analyses, excluding one study at a time and combining
the remaining studies. The results showed that, under all mentioned genetic models, no single study had an excess effect on
any pooled effect.
Publication bias evaluation
After excluding the key factors of heterogeneity between studies, no publication bias regarding the association between polymorphisms and overweight and obesity was detected in any of
the abovementioned genetic models using funnel plots (Figure 8).
Table 1: Relevant features of the studies included in the meta-analysis.
| Study |
Country |
Year |
Ethnicity |
Age |
Case |
Control |
NOS score |
| DD |
DI |
II |
DD |
DI |
II |
| Dalgaard et al. [6] |
Denmark |
1999 |
Caucasian |
adults |
371 |
293 |
80 |
432 |
364 |
76 |
8 |
| Evans et al. [7] |
Germany |
2000 |
Caucasian |
adults |
145 |
130 |
30 |
286 |
198 |
24 |
6 |
| Esterbauer et al. [8] |
Austria |
2001 |
Caucasian |
adults |
179 |
132 |
29 |
117 |
115 |
24 |
8 |
| Nieters et al. [9] |
Germany |
2002 |
Caucasian |
adults |
88 |
58 |
8 |
78 |
57 |
16 |
8 |
| Maestrini et al. [10] |
Italy |
2003 |
Caucasian |
adults |
211 |
124 |
25 |
51 |
42 |
10 |
7 |
| Feng et al. [11] |
China |
2004 |
Asian |
adults |
75 |
27 |
1 |
138 |
31 |
2 |
7 |
| Marti et al. [12] |
Spain |
2004 |
Caucasian |
adults |
83 |
63 |
11 |
92 |
52 |
6 |
7 |
| Berentzen et al. [13] |
Denmark |
2005 |
Caucasian |
adults |
277 |
227 |
64 |
356 |
294 |
67 |
8 |
| Hong et al. [14] |
China |
2005 |
Asian |
children |
165 |
35 |
1 |
190 |
36 |
4 |
8 |
| Ochoa et al. [15] |
Spain |
2007 |
Caucasian |
children |
103 |
71 |
18 |
79 |
76 |
11 |
7 |
| Kring et al. [16] |
Denmark |
2008 |
Caucasian |
adults |
104 |
94 |
24 |
166 |
127 |
21 |
8 |
| Liu et al. [17] |
China |
2012 |
Asian |
adults |
463 |
141 |
12 |
696 |
199 |
15 |
7 |
| Papazoglou et al. [18] |
Greece |
2012 |
Caucasian |
adults |
96 |
55 |
7 |
60 |
27 |
4 |
8 |
| Csernus et al. [19] |
Hungary |
2013 |
Caucasian |
children |
338 |
303 |
68 |
339 |
251 |
47 |
7 |
| Oguzkan et al. [20] |
turkey |
2013 |
Caucasian |
children |
46 |
31 |
23 |
65 |
30 |
5 |
7 |
| Say et al. [21] |
Malaysia |
2014 |
Asian |
adults |
181 |
77 |
7 |
505 |
145 |
11 |
7 |
| Gul et al. [22] |
Turkey |
2017 |
Caucasian |
children |
151 |
96 |
21 |
93 |
68 |
24 |
7 |
| Kaabi et al. [23] |
Saudi Arabia |
2018 |
Caucasian |
adults |
58 |
23 |
5 |
30 |
32 |
3 |
7 |
| Surniyantoro et al. [24] |
Indonesia |
2018 |
Asian |
adults |
68 |
25 |
7 |
68 |
29 |
3 |
7 |
| Verdi et al. [25] |
Turkey |
2020 |
Caucasian |
children |
72 |
29 |
1 |
53 |
29 |
5 |
6 |
Table 2: Pooled measures for the associations of the UCP2 45bp Ins/Del gene polymorphism with overweight and obesity.
| Population |
Inherited model |
Numbers of |
Pooled OR (95% CI) |
I2(%) |
| cases/controls |
FEM |
P value |
REM |
P value |
|
| Overall |
Allele |
11500/12948 |
1.08 (1.02, 1.15) |
0.011 |
1.06 (0.95, 1.19) |
0.296 |
67.5 |
| Dominant |
5750/6474 |
1.07 (0.99, 1.15) |
0.096 |
1.05 (0.92, 1.19) |
0.484 |
59.1 |
| Recessive |
5750/6474 |
1.24 (1.07, 1.43) |
0.004 |
1.21 (0.97, 1.53) |
0.098 |
46.3 |
| Heterozygote |
5308/6096 |
1.03 (0.96, 1.12) |
0.397 |
1.02 (0.91, 1.15) |
0.704 |
45.4 |
| Homozygote |
3716/4272 |
1.24 (1.06, 1.43) |
0.005 |
1.20 (0.93, 1.56) |
0.161 |
54.9 |
| Asian |
Allele |
2570/4144 |
1.18 (1.02, 1.36) |
0.027 |
1.18 (1.02, 1.37) |
0.025 |
0 |
| Dominant |
1285/2072 |
1.20 (1.02, 1.41) |
0.03 |
1.20 (1.01, 1.43) |
0.035 |
6.4 |
| Recessive |
1285/2072 |
1.27 (0.76, 2.11) |
0.358 |
1.30 (0.77, 2.19) |
0.32 |
0 |
| Heterozygote |
1257/2037 |
1.19 (1.01, 1.41) |
0.043 |
1.20 (0.99, 1.45) |
0.059 |
12.6 |
| Homozygote |
980/1632 |
1.31 (0.78, 2.18) |
0.303 |
1.35 (0.80, 2.27) |
0.261 |
0 |
| Caucasian |
Allele |
8930/8334 |
1.06 (0.99, 1.14) |
0.072 |
1.03 (0.89, 1.18) |
0.706 |
73.5 |
| Dominant |
4465/4402 |
1.03 (0.95, 1.12) |
0.448 |
1.00 (0.85, 1.16) |
0.971 |
64.6 |
| Recessive |
4465/4402 |
1.23 (1.06, 1.43) |
0.007 |
1.20 (0.92, 1.56) |
0.171 |
56.7 |
| Heterozygote |
4051/4059 |
1.00 (0.91, 1.09) |
0.914 |
0.97 (0.85, 1.11) |
0.672 |
47.8 |
| Homozygote |
2736/2640 |
1.23 (1.05, 1.44) |
0.009 |
1.18 (0.88, 1.59) |
0.275 |
64.1 |
| Adults |
Allele |
8356/10138 |
1.08 (1.01, 1.16) |
0.03 |
1.07 (0.95, 1.21) |
0.248 |
59.7 |
| Dominant |
4178/5069 |
1.06 (0.97, 1.16) |
0.164 |
1.06 (0.91, 1.22) |
0.461 |
58 |
| Recessive |
4178/5069 |
1.25 (1.02, 1.48) |
0.011 |
1.24 (1.02, 1.52) |
0.031 |
14.7 |
| Heterozygote |
3868/4787 |
1.03 (0.94, 1.13) |
0.51 |
1.03 (0.89, 1.19) |
0.691 |
52.1 |
| Homozygote |
2709/3357 |
1.24 (1.04, 1.48) |
0.017 |
1.23 (0.96, 1.57) |
0.1 |
34.6 |
| Children |
Allele |
3144/2810 |
1.08 (0.96, 1.22) |
0.189 |
1.04 (0.76, 1.42) |
0.795 |
81 |
| Dominant |
1572/1405 |
1.07 (0.92, 1.24) |
0.358 |
1.02 (0.76, 1.37) |
0.897 |
67.6 |
| Recessive |
1572/1405 |
1.20 (0.92, 1.58) |
0.181 |
1.09 (0.53, 2.23) |
0.81 |
75.1 |
| Heterozygote |
1440/1309 |
1.05 (0.89, 1.22) |
0.582 |
1.00 (0.81, 1.25) |
0.979 |
34.9 |
| Homozygote |
1007/915 |
1.23 (0.93, 1.63) |
0.152 |
1.06 (0.49, 2.30) |
0.874 |
77.5 |
Discussion
UCP2 plays an important role in the development and treatment of overweight and obesity, as an obesity candidate gene
involved in the regulation of glucose/lipid metabolism and energy homeostasis. UCP2 affects the susceptibility to obesity and
obesity-related diseases by decreasing the activity or expression
of these UCPs, thereby increasing oxidative phosphorylation coupling to reduce energy expenditure. Thus, the expression/activity levels of UCP2 influence the relationship between UCP2 gene
polymorphisms and obesity [26]. The 3'UTR region of the UCP2
Ins/Del polymorphic gene, which is only 158 bp away from the
transcription termination codon, may function by participating
in mRNA processing or transcriptional stability, and lower transcript stability may result in lower UCP2 protein translocation
[11]. These three polymorphisms are currently the most studied
in the UCP2 gene: 45bp Ins/Del in exon 8, missense variant in
exon 4 (rs660339, Ala55Val, C/T), and one in the promoter region
(rs659366, 866G/A) [12]. The association of one of these 45bp Ins/Del polymorphisms with overweight and obesity remains
highly controversial. Therefore, we systematically summarized 20
eligible case-control studies using the largest sample size available for this meta-analysis to assess the association between
UCP2 45bp Ins/Del with genetic susceptibility and overweight and
obesity and guide future studies.
The analysis showed that the UCP2 45bp Ins/Del gene polymorphism was significantly associated with genetic susceptibility
to overweight and obesity only in the recessive model (OR=1.24,
95%CI=1.07-1.43, P=0.004), but race-specific subgroup analysis
showed that the UCP2 45bp Ins/Del gene polymorphism was significantly associated with genetic susceptibility to overweight and
obesity in Asian populations in the allelic (OR=1.18, 95%CI=1.02-1.36, P=0.027), dominant (OR=1.20, 95%CI=1.02-1.41, P=0.030),
and heterozygote (OR=1.19, 95%CI=1.01-1.41, P=0.043) models,
and no significant association was found in Caucasian populations.
Brondani's study showed similar findings to this study that
polymorphism is significantly associated with increased BMI in
Asian populations [27]. However, in a study on Caucasian populations, the findings of Brondani et al. contradicted our findings
by suggesting that the UCP2 45bp Ins/Del gene polymorphism
was significantly associated with obesity in Caucasian populations
[28]. Comparative results from other studies have shown that individuals carrying the II genotype and Ins allele are at higher risk of
obesity than those carrying the DD genotype compared to other
genotypes or alleles [17,21,29]. Moreover, Ins allele carriers have
a higher BMI in some populations [30]. Conversely, Zhang et al.
and Surniyantoro et al. showed that 45bp Ins/Del gene polymorphism is not a risk factor for overweight and obesity [24,31]. The
nutritional characteristics of populations may influence the relationship between genetic variation and obesity, and food and cultural habits and environmental factors differ for each race, which
may influence the association between UCP polymorphisms and
obesity [24,32]. Therefore, future studies on obesity gene polymorphisms should consider environmental factors and dietary
habits. Moreover, there are numerous genes associated with obesity [33], and each population has different genetic variants in its
gene pool; these factors may also play a role in obesity. Kring et al.
concluded that there is a lack of significant correlation between
genetic variants and BMI because obesity is a mixed phenotype,
and several other proxies for overweight and obesity are available, such as waist circumference, waist circumference for a given
BMI, sagittal abdominal diameter, and waist-to-hip ratio [15],
which should be considered in future studies. Some researchers
have suggested that the effect of insertion polymorphisms may be
age-related and associated with late-onset obesity [8]. Therefore,
we conducted a subgroup analysis of the study population divided
into child and adult groups for age and only found significant associations in the recessive (OR=1.19, 95% CI=1.01-1.41, P=0.011)
and homozygote (OR=1.24, 95% CI=1.04-1.48, P=0.017) models in
the adult group, while the other groups did not show significant
associations. A study by Gul et al. concluded that UCP2 exon 8 Ins/
Del had no significant effect on the risk of obesity in adolescents,
which is similar to our findings, and that low HDL cholesterolemia may be associated with the Ins allele [12]. Gender-stratified
UCP2 45bp Ins/Del gene polymorphism analysis by Surniyantoro
et al. and Papazoglou et al. showed that gene polymorphisms
had opposite effects on male and female populations, with the
II genotype and I allele leading to reduced UCP2 expression and increased body weight in the male population. Conversely, in the
female population, the UCP2 45bp Ins/Del gene polymorphism is
recessively associated with obesity [22,24].
Heterogeneity between studies is a matter of concern, and high
heterogeneity was observed in all our analyses of genetic models
of the association of the UCP2 45bp Ins/Del gene polymorphism
with overweight and obesity. After stratifying the analysis by
ethnicity, heterogeneity was significantly lower in the analysis of
Asian populations; however, significant heterogeneity remained
in the analysis of Caucasian populations, suggesting some other
confounding factors in the study on Caucasian populations. The
sources of heterogeneity were worth exploring; therefore, a regression analysis was implemented, including covariates such as
age, sex, and sample size. However, none of these covariates individually or jointly explained the observed heterogeneity. Study
quality, general characteristics of participants, representation
of participants, gene-environment interactions, and genotyping
methods may contribute to heterogeneity. Without additional
information on the metabolic and clinical characteristics of the
articles analyzed, the role of these factors in the sources of heterogeneity is difficult to describe accurately.
Therefore, the results of the current meta-analysis should be
interpreted with caution. First, the definition of cases is not uniform across studies; in some studies, cases are defined as obese
populations, morbidly obese populations, or mixed populations of
overweight and obesity. Moreover, in some relevant studies, the
threshold values of overweight or obesity were different. Second,
due to technical limitations, we only retrieved studies published
in English and Chinese, which may lead to certain omissions. Lastly, our analysis revealed significant heterogeneity among studies
on Caucasian populations.
Conclusions
Our study showed a significant association between UCP2
45bp Ins/Del gene polymorphism and genetic susceptibility to
overweight and obesity in Asian populations but not in Caucasian
populations. Factors such as age, sex, environmental factors, and
dietary habits may influence the accuracy of this association. To
obtain strong evidence, a larger sample size and more adequate information are needed to conduct more detailed analytical studies.
Declarations
Acknowledgment: This research was funded by grants from
the National Natural Science Foundation of China (No. 82160578),
Natural Science Foundation of Jiangxi Province, China (No.
20212BCJ23024 and 20202BAB216029), Health Department of Jiangxi Province, China (No. 20198020), and Education Department
of Jiangxi Province, China (No. GJJ190019).
Conflict of interest: The authors have no conflict of interests.
References
- Trends in adult body-mass index in 200 countries from 1975 to 2014:
a pooled analysis of 1698 population-based measurement studies with 19•2 million participants. Lancet. 2016; 387: 1377-1396.
- Ng M, Fleming T, Robinson M, Thomson B, Graetz N, et al. Global,
regional, and national prevalence of overweight and obesity in
children and adults during 1980-2013: a systematic analysis for the Global Burden of Disease Study. Lancet. 2013; 384: 766-781.
- M. Blüher. Obesity: global epidemiology and pathogenesis. Nat
Rev Endocrinol. 2019; 15: 288-298.
- H. Oberkofler, Liu YM, Esterbauer H, Hell E, Krempler F, et al. Uncoupling protein-2 gene: reduced mRNA expression in intraperitoneal adipose tissue of obese humans. Diabetologia. 1998; 41:
940-946.
- Fleury C, Neverova M, Collins S, Raimbault S, Champigny O, et al.
Uncoupling protein-2: a novel gene linked to obesity and hyperinsulinemia. Nat Genet. 1997; 15: 269-272.
- Dalgaard LT, Sørensen TI, Andersen T, Hansen T, Pedersen O. An
untranslated insertion variant in the uncoupling protein 2 gene is
not related to body mass index and changes in body weight during
a 26-year follow-up in Danish Caucasian men. Diabetologia. 1999;
42: 1413-1416.
- D. Evans, Minouchehr S, Hagemann G, Mann WA, Wendt D, et al.
Frequency of and interaction between polymorphisms in the beta3-adrenergic receptor and in uncoupling proteins 1 and 2 and obesity
in Germans. Int J Obes Relat Metab Disord. 2000; 24: 1239-1245.
- H. Esterbauer, Schneitler C, Oberkofler H, Ebenbichler C, Paulweber B, et al. A common polymorphism in the promoter of UCP2 is
associated with decreased risk of obesity in middle-aged humans.
Nat Genet. 2001; 28: 178-183.
- Nieters, N. Becker, J. Linseisen. Polymorphisms in candidate obesity genes and their interaction with dietary intake of n-6 polyunsaturated fatty acids affect obesity risk in a sub-sample of the EPIC-Heidelberg cohort. Eur J Nutr. 2002; 41: 210-221.
- S. Maestrini, Podestà F, Di Blasio AM, Savia G, Brunani A, et al.
Lack of association between UCP2 gene polymorphisms and obesity phenotype in Italian Caucasians. J Endocrinol Invest. 2003; 26:
985-990.
- Marti A, Corbalán MS, Forga L, Martinez-González MA, Martinez
JA. Higher obesity risk associated with the exon-8 insertion of the
UCP2 gene in a Spanish case-control study. Nutrition. 2004; 20:
498-501.
- Qiwen F, Yan D, Qing S, Huili X, Jingnan Z. Association of uncoupling
protein 2 gene polymorphisms with type 2 diabetes in Chinese.
Journal of Shanghai Second Medical University. 2004; 108-110.
- Berentzen T, Dalgaard LT, Petersen L, Pedersen O, Sørensen TI.
Interactions between physical activity and variants of the genes
encoding uncoupling proteins -2 and -3 in relation to body weight
changes during a 10-y follow-up. Int J Obes (Lond). 2005; 29: 93-99.
- Qingrong H. Association of uncoupling protein 2 gene polymorphisms with obesity in children and adolescents. Chinese Journal
of Endocrinology and Metabolism. 2005; 545-546.
- Ochoa MC, Santos JL, Azcona C, Moreno-Aliaga MJ, Martínez-González MA, et al. Association between obesity and insulin resistance with UCP2-UCP3 gene variants in Spanish children and
adolescents. Mol Genet Metab. 2007; 92: 351-358.
- Kring SI, Larsen LH, Holst C, Toubro S, Hansen T, et al. Genotype-phenotype associations in obesity dependent on definition of the
obesity phenotype. Obes Facts. 2008; 1: 138-145.
- X. Liu, Zhang B, Liu X, Shen Y, Li J, et al. A 45-bp insertion/deletion
polymorphism in uncoupling protein 2 is not associated with obesity in a Chinese population. Biochem Genet. 2012; 50: 784-796.
- Papazoglou D, Papathanasiou P, Papanas N, Papatheodorou K,
Chatziangeli E, Nikitidis I, et al. Uncoupling protein-2 45-base pair
insertion/deletion polymorphism: is there an association with severe obesity and weight loss in morbidly obese subjects? Metab
Syndr Relat Disord. 2012; 10: 307-311.
- Csernus K, Pauler G, Erhardt E, Lanyi E, Molnar D. Uncoupling protein-2 gene polymorphisms are associated with obesity in Hungarian children. Acta Paediatr. 2013; 102: e200-204.
- Oguzkan-Balci S, Col-Araz N, Nacak M, Araz M, Sabanci H, et al.
Mitochondrial uncoupling protein 2 (UCP2) gene polymorphisms
are associated with childhood obesity and related metabolic disorders. J Pediatr Endocrinol Metab. 2013; 26: 277-283.
- Say YH, Ban ZL, Arumugam Y, Kaur T, Tan ML, et al. Uncoupling
protein 2 gene (UCP2) 45-bp I/D polymorphism is associated with
adiposity among Malaysian women. J Biosci. 2014; 39: 867-875.
- Gul, Ateş O, Özer S, Kasap T, Ensari E, et al. Role of the Polymorphisms of Uncoupling Protein Genes in Childhood Obesity and
Their Association with Obesity-Related Disturbances. Genet Test
Mol Biomarkers. 2017; 21: 531-538.
- Kaabi YA. The Deletion Polymorphism in Exon 8 of Uncoupling Protein 2 is Associated with Severe Obesity in a Saudi Arabian Case-control Study. Indian J Endocrinol Metab. 2018; 22: 200-203.
- Surniyantoro HNE, Sadewa AH, Hastuti P. Uncoupling Protein 2
(UCP2) as Genetic Risk Factor for Obesity in Indonesia is Different
in Gender Stratification. Kobe J Med Sci. 2018; 64: E64-e72.
- Verdi H, Kınık ST, Baysan-Çebi HP, Yalçın YY, Yazıcı-Güvercin AC, et
al. Uncoupling protein gene UCP1-3826A/G, UCP2 Ins/Del and
UCP3-55C/T polymorphisms in obese Turkish children. Turk J Pediatr. 2020; 62: 921-929.
- Azzu V, Brand MD. The on-off switches of the mitochondrial uncoupling proteins. Trends Biochem Sci. 2010; 35: 298-307.
- Brondani LA, Assmann TS, de Souza BM, Bouças AP, Canani LH, et
al. Meta-analysis reveals the association of common variants in
the uncoupling protein (UCP) 1-3 genes with body mass index variability. PLoS One. 2014; 9: e96411.
- Brondani LA, Assmann TS, de Souza BM, Bouças AP, Canani LH, et
al. Association of the UCP polymorphisms with susceptibility to
obesity: case-control study and meta-analysis. Mol Biol Rep. 2014;
41: 5053-5067.
- Hashemi M, Rezaei H, Kaykhaei MA, Taheri M. A 45-bp insertion/
deletion polymorphism of UCP2 gene is associated with metabolic
syndrome. J Diabetes Metab Disord. 2014; 13: 12.
- Jia JJ, Zhang X, Ge CR, Jois M. The polymorphisms of UCP2 and
UCP3 genes associated with fat metabolism, obesity and diabetes.
Obes Rev. 2009; 10: 519-526.
- Zhang M, Wang M, Zhao ZT. Uncoupling protein 2 gene polymorphisms in association with overweight and obesity susceptibility:
A meta-analysis. Meta Gene. 2014; 2: 143-159.
- Luan JA, Wong MY, Day NE, Wareham NJ. Sample size determination for studies of gene-environment interaction. Int J Epidemiol.
2001; 30: 1035-1040.
- Kim HJ, Lee SY, Kim CM. Association between gene polymorphisms
and obesity and physical fitness in Korean children. Biol Sport.
2018; 35: 21-27.